La vida [estructures i processos complexes]
- Introducció
- Parts i funcions: membrana i citoplasma, material genètic, ribosomes, mitocondris, cloroplasts. Les peces de la vida. Cicle de vida, mitosi
- Metabolisme: Catabolisme, fotosíntesi, glucòlisi, respiració. Anabolisme : biosíntesi, moviment, transport. Autoreplicació (Duplicació, morfogènesi, mitosi), transcripció. Codi genètic.
- Altres: la cèl·lula, com a arquitecte de la vida.
Introducció
[La cèl·lula en la seva versió més simple és com una bossa de plàstic, com a membrana, surant dins de l’aigua. A través de la membrana intercanvia nutrients [quins, oxígen, co2]. A dins hi té els “plànols” de DNA per generar noves proteïnes, proteïnes i enzims per fer el catabolisme i anabolisme, i eventualment, duplicar-se.
Els organismes pluricel·lulars som una “societat” de cèl·lules, pròpies i convidades que generem un entorn “aquàtic” adequat per a les nostres cèl·lules, com un aquari. Un aquari líquid en els organismes aquàtics, i un aquari sòlid pels terrestres.]

Teoria cel·lular:”la cèl·lula és una unitat morfològica de tot ésser viu; és a dir, que en els éssers vius tot està format per cèl·lules o pels seus productes de secreció.” No hi ha generació espontània, Omnis cellula ex cellula (Rudolf Virchow). Les funcions vitals dels organismes tenen lloc o bé dins de les cèl·lules, o en el seu entorn immediat. Cada cèl·lula és un sistema obert que intercanvia matèria i energia amb el seu ambient augmentant l’entropia de fora. Cada cèl·lula conté tota la informació hereditària necessària per al control del seu propi cicle i el desenvolupament i funcionament d’un organisme de la seva espècie, així com per la transmissió d’aquesta informació a la següent generació cel·lular.” (Ciències de la vida sXIX)
Parts i funcions
Característiques funcionals: Nutrició (agafar substàncies de l’ambient i transformar-les, obtenint components i energia i retornant els residus), Creixement i proliferació cel·lular, Diferenciació (els gens s’expressen segons diferents estímuls externs), Senyalització (resposta a estímuls químics i físics, comunicació amb altres cèl·lules, per senyals químics com hormones, o neurotransmissors). Evolució.
Mida [Tal com indica l’article New Yorker 7/3/2022 encara no podem observar què passa dins d’una cèl·lula, a un escala entre els àtoms de 1 å, 10-10m, i els bacteris de 2 μm, 10-6m de longitud. El mycoplasma genitalium és de 0.2 μm i el límit teòric seria de 0.17 μm. En l’organisme humà, els hematies fan 7 μm, els espermatozous 53 μm, els òvuls de 150 μm.
Membrana: Individus gràcies a una membrana que les separa i comunica amb l’exterior, que controla els moviments cel·lulars i que manté el potencial elèctric de la cèl·lula.
Dues capes de lípid (45%) que envolten unes proteïnes flotants (55%). Formen una barrera (¸10-8m) selectiva que deixa passar l’aigua i algunes substàncies (reconeixement per enzims) arribant a tenir un paper actiu en el transport [aliments i deixalles). La paret cel.lular, no sempre present, està formada per polisacàrids amb un gruix d’uns 20 nm i evita que la cèl·lula exploti quan no està limitada per altres. Deixa passar molècules petites. A la membrana hi ha també els enzims que converteixen els aliments en ATP, tasca que a les cèl·lules eucariotes acompliran els mitocondris.
Dutrochet (1827) descobreix el paper de l’òsmosi a la membrana i més tard Van’t Hoff i Arrhenius n’enuncien les lleis (1884). 1935 Danielli i Dawson indiquen que la membrana cel·lular oleica està recoberta de proteïnes. El 1972 Singer i Nicolson ho refinaran dient que estan intercalades.
Citoplasma: el que hi ha a l’interior de la membrana, diversos orgànuls en una una dissolució d’aigua, sals inorgàniques, proteïnes del metabolisme i intermediaris del procés metabòlic, el citosol. Pot tenir un citoesquelet amb microtúbuls i microfilaments que li aporta certa consistència i ordena l’espai.
Material genètic. Cromosomes [no em queda clar si sempre està organitzat en cromosomes o bé només ho fan en el moment de la divisió] , una doble hèlix de DNA de 2 nm de diàmetre i 1 mm de llarg enrotllat molt compacte.[108 elements!]. Servirà per transcriure proteïnes i autoreplicar-se. les ARN polimerases transcriuen ARN missatger contínuament, que, exportat al citosol, és traduït a proteïna segons les necessitats fisiològiques. A més, depenent del moment del cicle cel·lular, aquest ADN pot entrar en replicació com a pas previ a la mitosi. [Àcids nucleics]
A les cèl·lules eucariotes està separat de la resta del citoplasma per una membrana formant un nucli.
Ribosomes: boles de 18 nm en un nombre de 15000 a l’E.Coli (l’organisme més ben conegut) on els RNA copiat del DNA codifiquen la síntesi de proteïnes. [es copia una part dels plànols del DNA, al mRNA i es du al “taller” on es construirà la proteïna]. Poden estar agrupats al reticle endoplasmàtic. Els processos de les proteïnes són facilitats per l’estructura de l’aparell de Golgi.
El m-RNA, RNA missatger, copia el “negatiu” a partir del DNA. Es troba amb un t-RNA que ha acoblat l’aminoàcid corresponent. [Si ho entenc bé, el m-RNA tindria la longitud d’un gen, i el t-RNA només un codó per un aminoàcid]

Grànuls de reserva: polímers de sucres, reserves d’energia per a l’organisme.
Mitocondris: orgànuls encarregats de l’obtenció d’energia a través de la respiració cel·lular. Intervenen en el cicle de Krebs, la fosforilació oxidativa i la cadena de transport d’electrons de la respiració. S’haurien assimilat de bacteris per endosimbiosi i tenen material genètic independent.

Als eucariotes vegetals hi trobem:
- Cloroplasts: Orgànuls de color verd que duen a terme la fotosíntesi [només són a les fulles? no a les arrels?]. Segons la hipòtesi de la simbiosi s’haurien integrat a partir de cianobacteris i tenen material genètic independent.
- Vacuoles: acumulen reserves, faciliten l’intercanvi amb l’exterior i mantenen la turgència de la cèl·lula.
Als eucariotes animals hi trobem liposomes, enzims hidrolítics que serveixen per degradar molècules.
Les peces de la vida
[Si volem fer una estimació grollera de quantes i quines peces formen la vida tenim, els 4 nucleòtids del DNA, les bases adenina (A), timina (T), citosina (C) i guanina (G), cadascuna formada per uns 10 àtoms, i els 20 aminoàcids que formen les proteïnes.] Bioquímica.
Al bacteri E. coli hi ha uns 4300 gens que codifiquen proteïnes. Les proteïnes tenen uns 300 aminoàcids de mitjana. Així tindríem 1.29 106 aminoàcids. Amb uns 12 àtoms per aminoàcids tenim 2.3 107 àtoms de proteïnes. [no sabem quantes còpies de cada proteïna, però]. En els humans hi hauria de 20.000 a 25.000 proteïnes, entre 300 i 10000 aminoàcids. Així que en una cèl·lula hi hauria 22.000×2.000 uns 4.4 107 aminoàcids, amb uns 5 108 àtoms de proteïnes.
A l’E. coli hi ha 4.6M de parelles de nucleòtids que donarien uns 9.2 107 àtoms. En una cèl·lula humana hi ha uns 3.000 M de parelles de bases, que serien uns 6 1010 àtoms.
Les cèl·lules vives tindrien de l’ordre de 109 parelles de bases al DNA i unes 10.000 proteïnes amb uns 2000 aminoàcids, 2 107 en total.
Procariotes: (bacteris i arquebacteris) les cèl·lules no tenen estructura interior ni nucli definit, tots els orgànuls estan dispersos al citosol.
Eucariotes animals
Diàmetre 20µm (0.002 mm) En un cm3 n’hi ha 106 deixant espai intercelular i tot. (Lehninger p.35, Atlas pp.23-51).
La membrana té més proporció de lípids que els procariotes.
El nucli fa uns 5 nm diàmetre. La doble hèlix del DNA s’enrotlla sobre proteïnes histones formant un filament de cromatina. Aquests s’agrupen en cromosomes, bastonets d’uns 0,0002 mm que es dividiran en la divisió nuclear. El número de cromosomes és fix per cada espècie, són dobles en les cèl·lules somàtiques (diploides) i simples en les c.sexuals (haploides). Els nucleols són concentracions de RNA. [el material genètic adopta la forma de cromosoma en la fase de mitosi del cicle de vida).
Els mitocondris produeixen energia oxidant glúcids. Origen per endosimbiosi. N’hi ha uns 500, 20% citoplasma, elipsoides de 1µm d. plens d’enzims on els glúcids, lípids i aminoàcids més O2 s’oxiden a CO2 i H2O alliberant energia que serveix per formar ATP.
Complex de Golgi, un reticle de cossos buits que aïlla certes substàncies del citoplasma tot expulsant-les a l’exterior. Serà important en les cèl·lules especialitzades en secreció. Peroxisomes: Vesícules amb enzims d’oxidació. Lisosomes: vesícules amb enzims (fosfatasa àcida) que digereixen substàncies absorbides o les deixalles.
Reticle endoplasmàtic, xarxa de conductes de superfície rugosa on hi ha els ribosomes que duen a terme la síntesi proteica.

Eucariotes vegetals
Com la procariota, membrana, citoplasma, nucli (separat), mitocondries, reticle endoplasmàtic, i pròpiament vegetal, cloroplastos i vacuoles. (Lehninger p.37). Cloroplast, receptors d’energia lluminosa que converteixen en ATP alliberant oxígen i consumint CO2 i H2O. Vacuoles, orgànuls que van absorbint restes dels processos, expulsant deixalles i augmentant de volum al llarg de la vida de la c. Contenen sucres dissolts, sals, proteïnes.

Cicle de vida
Interfase [cèl·lula nova després d’una mitosi]
- G1: Creixement cel·lular amb síntesi de proteïnes i d’ARN. transcripció i reactivació del mRNA, concentració de nucleòtids.
- S: s duplica l’hèlix de DNA, es produeixen histones. cada cromosoma es duplica i queda format per dues cromàtides idèntiques.
- G2: Continua la síntesi de proteïnes i ARN. Es fon el reticle endoplasmàtic, la cèl. perd contecte amb les veïnes i s’arrodoneix. S’acaba quan els cromosomes comencen a condensar-se al principi de la mitosi.
Mitosi (Atlas p.39)
- Profase: la cromatina [el material genètic] es concentra en cromosomes. Prometafase: es desfà la membrana del nucli i entren microtúbuls que desplacen els centrosomes.
- Metafase: S’acaben d’espiralitzar les cromàtides, els cromosomes s’alineen entre els dos centrosomes.
- Anafase: els cromosomes de separen i es desplacen, , se separen les cromàtides germanes dirigint-se als pols.
- Telofase i citokinesi: Els cromosomes tornen a disposar-se normalment i es dissolen en cromatina. A partir del reticle endoplasmàtic es torna a formar l’envoltura nuclear entorn de cada centròmer. A l’equador es genera una biomembrana que separarà les dues noves cèl·lules.
[Quin és el rellotge que indica el ritme de reproducció i mort?]

(En el procés de meiosi de la reproducció a la metafase es fusionen les cromàtides del pare i la mare (2N →N), a l’anafase se separen (N→2N) i a la telofase es divideixen (2N). Després a la meiosi II es tornen a dividir i tindrem 4 gàmetes haploides.)
[Procés general de reproducció, creixement i mort, setmanes, mesos, anys? El cos en constant reconstrucció] [ Procés cíclic d’assimilació de nutrients i biosíntesi, hores]
[Manca una explicació de la degradació de la cèl·lula, la mort]

Metabolisme
[igual que explotem la terra per obtenir energia i materials, l’organisme fa com una “mineria” del medi, ingerint nutrients i respirant oxígen. Amb el que “excavem” (1) carreguem les nostres bateries de ATP, (2) preparem materials i reconstruïm tot el que es va desfent (3) i tenim un circuit d’eliminació de deixalles]
Catabolisme: Reaccions químiques per obtenir components i energia. La ruta catabòlica degrada les grans molècules ingerides (proteïnes, polisacàrids i lípids) a components (aminoàcids, pentoses i àcids grassos), a acetil-CoA i finalment a NH3, H2O i CO2.
Anabolisme: biosíntesi, transport i contracció muscular. La ruta anabòlica parteix d’aquí per sintetitzar també en tres fases, els components superiors. La ruta catabòlica descendent i exergònica convergent a elements simples no passa pels mateixos passos intermitjos que l’anabòlica que puja i es divergeix en components superiors especialitzats. Això es deu a causes energètiques. Són regulades per diferents enzims (Lehninger p.381).
Cicle del carboni: Autòtrofs (llum) i heteròtrofs (química) es complementen mútuament formant un cicle on el CO2 es torna glucosa amb energia lumínica (plantes) que després és cremada en CO2 altra vegada amb la respiració animal [i vegetal] dissipant calor.
Cicle del Nitrogen: El gas nitrogen N2 és molt inert i només pot ser fixat per bacteris que el deixen com a amoníac o Nitrat → (Plantes superiors) → aminoàcids → (animals superiors) → amoníac/urea → (bacteris nitrificants, nitrosomes) → Nitrit (B.Nitrificants, nitrobacter) → Nitrat. [els grans cicles]
Catabolisme
Hidrats de carboni: Obtenció d’energia a partir de la glucosa [que es genera a les plantes per fotosíntesi] per glucòlisi i respiració cel·lular (i fermentació en alguns bacteris).
Greixos: trencament dels triglicèrids en àcids grassos i glicerol, mitjançant la incorporació de tres molècules d’aigua i l’ajuda d’enzims anomenades lipases. Els àcids grassos, guardats en forma de triglicèrids, són una reserva d’energia més concentrada que els sucres (9 kcal/g contra 4 kcal) i es fan servir pels animals superiors en cassos de dejuni o hibernació. Són esterificats fins a formar un acetil-CoA i després carreguen ATP a les mitocondries. Un exemple és: àcid palmític + 23O2 + 129Pi + 129 ADP = 16CO2 + 145 H2O + 129 ATP
procés on es recupera en ATP un 40% de l’energia de l’àcid.
Proteïnes: Escissió de les cadenes polipeptídiques en els seus aminoàcids mitjançant enzims anomenats proteases. Els aminoàcids constitueixen una altra font d’energia en els vertebrats. Les proteïnes són descomposades en el tub digestiu (tripsina) i passen a la sang per on seran conduïdes al fetge on tindrà lloc el seu catabolisme. Una degradació oxidativa els deixa en Acetil-CoA amb amoníac com a subproducte (asparagina+H2O -> Aspartato+ NH3). L’amoníac s’excreta com a urea, format-se així un cicle del nitrogen (cicle de la urea, Krebs 1930). La síntesi de la urea consumeix quatre grups fosfat:
2NH3+CO2+3ATP+3H2O -> urea + 2ADP + AMP + 4Pi

(0) Fotosíntesi: llum + CO2 → sucres

Als cloroplasts de les cèl·lules de les fulles té lloc la fotosíntesi, que en resum és:
6 CO₂ + 12 H₂O + llum → C₆H₁₂O₆ + 6 O₂ + 6 H₂O
Cicle de Calvin (1961)

Garreu (1849) mostra que l’evaporació d’aigua a les fulles és proporcional al nombre d’estomes. Liebig mostrarà que l’humus i els fems no aporten altra cosa que minerals. Sachs i altres determinaran els que són essencials: nitrogen, fòsfor, sofre, calci, potassi i magnesi. A més tenen un paper en petites quantitats, el ferro, zenc, magnanesi, i boro. Juntament amb l’aigua, carboni i oxigen són els 13 elements que componen les plantes.
En el cas dels bacteris sulfurats el donador d’electrons és sofre en lloc d’oxígen, la reacció general seria 2H2D + CO2 = (CH2O) + H2O + 2D, on D és el donador d’electrons.
La meitat de la fotosíntesi que es fa a la terra es duu a terme per plantes superiors i l’altra per les algues microscòpiques dels oceans, diatomees i dinoflagelats. Un m2 de fulla origina 1g sucre per hora. Es calcula que s’aprofita un 0.12% de la radiació incident. La reacció té lloc als cloroplasts que contenen els pigments capaços d’absorbir la llum ( Lehninger p.608-610). Els pigments presenten un màxim d’absorció per llum de 700 nm i 680 nm.
Fase lluminosa: [carreguem les piles ADP a ATP] Dos fotosistemes es complementen per incorporar els e- de l’oxígen de l’aigua (contra la tendència natural redox) a un enzim NADP alhora que carreguen ADP a ATP:
2 H2O + 2 NADP+ + 3 ADP + 3 Pi + llum → 2 NADPH + 2 H+ + 3 ATP + O2
La fase fosca: [fem servir la pila que obtenir sucre a partir de CO2]
3 CO₂ + 9 ATP + 6 NADPH + 6 H+ → C₃H₆O₃-fosfat + 9 ADP + 8 Pi + 6 NADP+ + 3 H₂O
Respiració aeròbica

C1 indica CO₂, C6 indica 6 àtoms de carboni, glucosa
El procés es resumeix (en el cas que el sucre sigui la glucosa) com:
C₆ H₁₂ O₆ + 6O₂ → 6CO₂ + 6H₂O + energia (ATP) ΔG = −2880 kJ per mol of C6H12O6
i té 4 etapes (és regulat per la càrrega energètica, és a dir, és estimulada per la concentració de ADP o AMP i inhibida per la de ATP.)
- (1) Glucòlisi: sucres → Piruvat: glucosa + 2 NAD+ + 2 ADP + 2 Pi → 2 piruvat + 2 NADH + 2 ATP + 2H+ + H₂O. Té lloc al citosol de la cèl·lula. Ho duen a terme totes les cèl·lules vives. glucosa + 2ADP + 2Pi -> àcid làctic + 2ATP + 2H₂O, glucosa + 2ADP + 2Pi -> 2etanol + 2CO₂ + 2ATP + 2H₂O
- (2) Descarboxilació oxidativa : Piruvat → Acetil-CoA Té lloc als mitocondris. El piruvat és oxidat després a Acetil-CoA. (Lehninger p.450)
Àcid pirúvic + NAD + HSCoA → Acetil-CoA + NADH + H+ + CO₂ - (3) Cicle de Krebs (1937) Un conjunt de 9 reaccions. Per cada glucosa es fan dues voltes. L’acetil-CoA és acabat d’oxidar pels processos següents (Atlas p.303) Té lloc a les mitocondries i segueix les etapes: CH3CO-S-CoA -> Acetil-CoA, Citrat, Isocitrat, Ó-Oxoglutarat, Succinil-CoA, Succinat, Fumarat, Malat, Oxalacetat, Acetil-CoA. A cada volta s’incorpora una molècula d’àcid acètic, se n’alliberen dos de CO2 i 8 H+ i 8 e- que seran usats en el procés posterior (Lehninger p.456). Els àcids intermitjos es recuperen llevat de quan els productes intermitjos són aprofitats per a la biosíntesi. Aleshores cal reposar-los amb reaccions laterals. [ Surt NADH i H+ que en la següent fase es combina amb oxígen per donar aigua i es recupera el NADH]

- (4) Cadena respiratòria i Fosforilació oxidativa. També té lloc als mitocondris. [Aquí és on entra l’oxígen per acceptar electrons i el 2H+ O- es convertiran en aigua, aquí hi hauria la combustió que suggeria Lavoisier] Es reoxiden els coenzims que que s’han reduït en les etapes anteriors i es produeix energia en forma de ATP. Els electrons i els protons implicats en aquests processos són cedits definitivament l’O₂ que es redueix a aigua.
Transport Electrònic (Lehninger p.505) Els electrons alliberats en el cicle anterior són transportats pels enzims NADH i NADPH en reaccions del tipus: substrat.reduït + NAD+ <-> s.oxidat + NADH + H+, i posteriorment a deshidrogenases, proteïnes ferro sulfurades i citocroms. Cada etapa és una reacció redox amb uns potencials estàndar de l’ordre de 0.5V (Len.p.490). En aquest procés de transferència electrònica s’allibera molta energia lliure: AG=nFAE (n núm. e- transferits, F=23.000 cal, AE diferència de potencials redox) ¸ -52.7 kcal/mol [l’hidrogen s’oxida a aigua cedint e- a l’oxígen atmosfèric. La gran energia perduda s’usa per ADP -> ATP]
Fosforilació oxidativa es duu a terme amb l’energia proporcionada pel transport electrònic: ADP+Pi+(2H++1/2O2)=ATP+H2O + 7kcal/mol. Aquest procés demana uns gradients de concentració d’ions que tenen lloc a les membranes dels mitocondris.
Alguns bacteris poden obtenir energia sense oxígen, amb respiració anaeròbica: 10H+ + sulfat ( SO24) + 8e- → H2S + 4 H₂O. És 15 vegades menys eficient que la respiració aeròbica. [no veig on entra la glucosa]
La fermentació, descoberta per Pasteur, no hi ha transferència d’electrons .
Anabolisme
Els components i energia obtinguda del catabolisme servirà per:
- Treball químic, biosíntesi. En general la ruta de la síntesi és diferent a la de la degradació catabòlica. La síntesi és possible gràcies a l’acoblament amb reaccions que hidrolitzen l’ATP subministrant l’energia per convertir en endergònic un procés exergònic. La ruta anabòlica està regulada per la concentració del producte final (que actúa sobre l’enzim de la primera etapa de manera que el procés mai s’interromp a la meitat). L’economia dels processos cel·lulars millora ja que només se sintetitza si manca producte, aleshores es consumeix ATP i es desencadena el procés catabòlic just per compensar la pèrdua.
- Treball mecànic, contracció, divisió cel·lular.
- Treball osmòtic o de concentració, transport a membranes.
- (Treball elèctric i tèrmic)
BIOSÍNTESI
- Biosíntesi de glúcids i lípids
Glúcids. (L. 23) és el procés més important de la bioesfera. Als autòtrofs és: CO₂ → hexoses→ midó (reserva) i cel.lulosa (parets) a les plantes. Als heteròtrofs: piruvat o lactat o aminoàcids → glucosa → glucogen (quitina insectes). El procés comença als mitocondris i acaba al citosol. Als vertebrats es fa al fetge.
Lípids. (L. 24) són un mitjà de guardar energia de manera més concentrada que els sucres i també tenen un paper regulador (prostaglandines). A partir del Acetil-CoA i) Acids grassos → Prostaglandines i triglicèrids, ii) Hidroximetilglutaril-CoA → (terperns i cetones) Colesterol → àcids biliars, esterols fecals, hormones esteroides. Els procés té lloc als citosols a partir de l’acetil-CoA resultant de la respiració mitocondrial. En els vertebrats es fa al fetge, teixit adipós i glàndules mamàries. - Biosíntesi d’aminoàcids i proteïnes
(L.25) Alguns organismes són capaços de fixar directament el nitrogen atmosfèric (bacteris i les plantes lleguminoses que els contenen). Totes les plantes són capaces de sintetitzar els 20 aminoàcids precursors de les proteïnes a partir d’amoníacs, nitrits (N2O3) o nitrats (N2O5) mentre que alguns animals superiors com l’home o la rata només en poden sintetitzar 10, havent d’ingerir els altres. Tots els aminoàcids es formen a partir de subproductes de processos catabòlics com la glucòlisi o el cicle de Krebs.
De la glucòlisi la 6fosfat-glucosa és precursor de la histidina, el 3fosfoglicerat és precursor de la glycine i cysteine; phosphoenol pyruvate + , 3-phosphoglycerate-derivative erythrose 4-phosphate forma tryptophan, phenylalanine, and tyrosine.
Del cicle de Krebs α-ketoglutarate és convertit en glutamate i després glutamine, proline, and arginine. L’oxaloacetate passa a aspartate i després a asparagine, methionine, threonine, and lysine.
La síntesi dels no essencials fa oxoglutarat → àcid glutàmic → glutamina i prolina, piruvat i oxalacetat → alanina i àcid aspàrtic, fenilalanina → tirosina, metionina → cisteïna, fosfoglicerat → serina → glicina. Els aminoàcids essencials són: → àcid aspàrtic → treonina i metionina, piruvats → valina, isoleucina, leucina, àcid glutàmic → ornitina → arginina, → histidina, àcid pirúvic → aromàtics com fenilalanina i triptòfan.
El pas d’aminoàcids a proteïnes és tan complex que ha de ser codificat amb molècules d’informació com és el RNA. [té lloc als ribosomes] - Biosíntesi de nucleòtids
(L. 26) La poden fer gairebé tots els organismes. Fosfo-D-ribosa+ … + 2 anells → purines → adenina i guanina (AMP i GMP). Acid oròtic + ribosa → uracil i citosina (UMP i CTP). Els ribonucleòsids s’obtenen per reducció dels Desoxir.
La síntesi dels àcids nucleics demana la intervenció de molècules d’informació, del codi genètic.
MOVIMENT
Conversió d’energia química ATP a energia mecànica per desplaçament de filament de proteïna miosina sobre els d’actina. (Tubulina) L’ATP es converteix en energia mecànica mitjantçant la contracció de filaments. Aquesta contracció és la base de i) moviment muscular (normalment consumeix un 30% de l’ATP produït, arribant al 85% en una cursa de sprint, amb tensions de 3.5 kg/cm2 arribant a 30 tones en tot el cos!) ii) organització del contingut celular iii) divisió cel. iv) activitat de cilis i flagelos v) transport de material.
- Moviment muscular
Miofibrilles, conjunt de sarcòmers miosina i actina. Un múscul està format per miofibrilles amb uns segments, els sarcòmers, que poden ser de dues menes, gruixuts i anisòtrops o prims i isòtrops. Cada segment està format per un tipus de filament, els prims d’actina i els gruixuts de miosina que es poden desplaçar al llarg dels anteriors. La longitud de cada filament es conserva, només varia la posició relativa (L. p.761). La proteïna actina, globular, s’uneix formant cadenes d’unes 150 peces que s’enrotllen format hèlixs dobles (filament prim). Cada 7 unitats hi ha una molècula de troponina que pot fixar el ió Ca2+. La molècula de miosina és una molècula molt llarga (160 nm) amb un cap globular. La miosina forma un complex amb l’ATP i quan hi ha l’impuls nerviós l’hidrolitza tot activant el cap que formarà una unió amb el segment molecular més proper. Aquestes molècules són al mig d’un sarcoplasma i un reticle sarcoplasmàtic entre el qual s’intercanvien Ca+. L’impuls nerviós introdueix Ca del reticle al sarcoplasma. El Ca es fixa a la troponina, en varia la configuració i lliga la miosina tot produïnt-se la contracció. El retorn del Ca del sarcoplasma al reticle allibera els ponts relaxant el muscle. - Cilis, flagels
Conjunt de microtúbuls formats per tubulina, una proteïna de forma allargada que en grups de 10-14 forma un microtúbul de 10-5 mm de diàmetre i 10-2 de llarg. Els microtúbuls donen rigidesa a les regions plasmàtiques. Grups d’uns 27 microtúbuls formen centriols que intervenen en la divisió celular. Varis microtúbuls es poden unir per formar un cili (nombrosos i curts) o un flagel (2 o 3 per cel. i llargs). L’energia de l’ATP serveix per desencadenar una variació dels angles de segments correlatius que resulta en un moviment de fuet (Atlas p.17). - Transport
Desplaçament de molècules a favor o contra un gradient de concentració. Passiu si és a favor d’un gradient de concentració i actiu quan a través d’una membrana es força l’entrada de solut contra el gradient. A la membrana hi ha enzims transportadors que s’acoblen al substrat que per si sol no la pot traspassar, i el fan entrar tot consumint ATP. Així ho fa la Na+K+ATPasa amb els ions K+, la concentració dels quals intervé en processos de glucòlisi, ribosomes i senyals nerviosos. En els mamífers es manté una concentració alta de K+ i baixa de Na+ dins la cèl·lula mentre que el fluid extracel·lular té concentracions inverses. (L. 28, Atlas p.25).
Autoreplicació i transcripció, DNA i RNA
Síntesi d’una cadena de DNA per aparellament de nucleòtids complementaris.(Lehninger 31 i 32)(Atlas p.33s) [DNA a química orgànica]. El pes molecular del DNA dels virus més petis és de 106, el dels bacteris de 109 i el dels cromosomes eucariòtics de 80 109 [Hi ha més diferència entre un virus i un bacteri, 103 que entre un bacteri i l’home, 80. Comptant que cada nucleòtid pesa 102 i que la cadena és doble, tenim ¸ 108 unitats d’informació, cabria en un disc de 100 Megabytes!].En els virus el DNA és lineal enrotllat, als bacteris és un llaç continu, a les cèl·lules eucariotes està associat a proteïnes com la histona formant la cromatina.
- Replicació. El mecanisme (Meselson-Stahl 1958) és semiconservador: la doble hèlix es desenrotlla i cada una forma la seva complementària, de manera que el nou DNA està format per un bri vell i un de nou (el mecanisme conservador, sense desenrotllar-se, produiria un DNA fill totalment nou). El procés comença quan la RNA-polimerasa identifica els punts d’iniciació, trenquen la doble hèlix i sintetitzen 500 unitats de RNA com a cebador [iniciador, desencadenant], la DNA polimerasa Ó segueix a partir del RNA inicial afegint DNA (120-1000 unitats), la DNA polimerasa ß acaba la feina ( avegades intervé una gamma) i finalment s’elimina el RNA inicial. El nou bri té una polaritat complementària [les parts 3’i 5′ de la pentosa estan oposades]. La DNA-ligasa lliga els dos brins complementaris amb ponts d’hidrogen formant la doble hèlix.
- Morfogènesi. Estructura 3D de l’hèlix de DNA, cromosoma. (L. 36) L’autocatàlisi del DNA explica la duplicació d’una seqüència lineal de nucleòtids o aminoàcids (→ codi genètic). Cóm es forma l’estructura 3D del DNA i el cromosoma? cóm es formen les parts 3D com els complexos enzimàtics, les membranes i orgànuls i les diverses estructures supracelulars? El plegament es deu a minimitzar l’energia lliure. La mateixa raó explica l’associació de diferents molècules sense que arribin a formar enllaços covalents. Així es formen capes, fils. (S’ha estudiat en estructures simples com els virus o ribosomes).
- Mitosi. (veure més amunt).Duplicació celular
Transcripció i Síntesi de Proteïnes
B1420.1.DP Un gen és transportat pel mRNA, s’aparella amb tRNA que té 3 nucleòtids que s’aparellaran amb un aminoàcid d’acord amb el codi genètic. Síntesi de proteïnes regulada per gens repressors fixats a l’operon.
Les RNA-polimerases(-DNA-dirigides) transcriuen tres menes de RNA a partir del codi de DNA (eines per a la síntesi de proteïnes):
rRNA: RNA ribosòmic, de paper encara sense esclarir del tot, possiblement amb funcions catalítiques, RNA-pol I. Fixa el mRNA.
mRNA: RNA missatger, per la RNA-pol II, transporta una seqüència de codi. Hi ha uns punts d’iniciació i de final.
tRNA: RNA de transferència, per la RNA-pol III, és una estructura en forma de trèbol, amb un dels braços anticodó (3 nucleòtids) que encaixa amb un codó (3n) del mRNA, i un altre braç aceptor amb capacitat de fixar aminoàcids.
Codi genètic
Cada grup de tres nucleòtids ( codó) correspon a un aminoàcid (hi ha redundància de 64=43 codons contra 20 aminoàcids):
UUU Phe UCU Ser UAU Tyr UGU Cys
UUC Phe UCC Ser UAC Tyr UGC Cys
UUA Leu UCA Ser UAA End UGA End
UUG Leu UCG Ser UAG End UGG Trp
CUU Leu CCU Pro CAU His CGU Arg
CUC Leu CCC Pro CAC His CGC Arg
CUA Leu CCA Pro CAA Gln CGA Arg
CUG Leu CCG Pro CAG Gln CGG Arg
AUU Ile ACU Thr AAU Asn AGU Ser
AUC Ile ACC Thr AAC Asn AGC Ser
AUA Ile ACA Thr AAA Lys AGA Arg
AUG Met ACG Thr AAG Lys AGG Arg
GUU Val GCU Ala GAU Asp GGU Gly
GUC Val GCC Ala GAC Asp GGC Gly
GUA Val GCA Ala GAA Glu GGA Gly
GUG Val GCG Ala GAG Glu GGG Gly
[El mRNA es trasllada al ribosoma?] Els aminoàcids es fixen al t-RNA per acció d’uns grans enzims (pes molecular 150.000), les sintetases de l’aminoacil-tRNA, tot consumint ATP. Els anticodons del t-RNA van encaixant a la seqüència del m-RNA alhora que un altre enzim va separant els aminoàcids ja enllaçats del t-RNA. Arribats al final de la seqüència se separa el darrer tRNA, el mRNA i el polipèptid acabat de formar. El conjunt de DNA que correspon a una proteïna determinada (que transcriu una seqüència de mRNA) es diu gen.
El codi genètic és universal, comú a tots els sers vius. El “dogma central de la biologia molecular” (Crick 1958) afirma que la informació sempre flueix del polinucleòtid al polipèptid i no a l’inrevés. L’aparellament codó-anticodó i l’assignació codó-aminoàcid no té una explicació en termes d’energia d’enllaç que justifiqui el codi universal. El codi funciona gràcies a l’existència dels enzims que guien el procés. I aquests enzims existeixen gràcies a estar codificats en DNA mitjantçant el codi universal. Es a dir que potser el codi no és únic però un cop se n’ha format un per atzar entre diverses fluctuacions, l’avantatge selectiu que té és tan gran que és el que es reprodueix més ràpid, tot consumint la majoria de recursos i eliminant els competidors (Hipercicles Eigen 1971).
La meitat de les proteïnes sintetitzades és estructural i l’altra meitat enzimàtica.
Regulació de l’expressió genètica (L. 35)(A.467) Als procariotes, quan certs productes ja no són necessaris per a la cèl.lula, la producció del conjunt d’enzims que els genera és reprimit, (o estimulat e cas contrari). El conjunt de gens corresponent a aquest grup d’enzims s’anomena operon. Un operon conté els gens estructurals codificadors dels enzims, un gen regulador que codifica la proteïna repressora i un operador units entre sí. Si hi ha substrat pels enzims (→ regulació de l’anabolisme), aquest s’uneix a la proteïna repressora tot alliberant l’operador (Model de Jacob-Monod, 1961). La producció d’enzims es pot regular segons el medi, de manera que en unes condicions el repressor és fixat a l’operon inhibint la síntesi.
Als eucariotes la regulació és molt més complexa tot intervenint les hormones a més de la presència dels substrats dels enzims. (A.471 → paper de les histones, model de Britton-Davidson).
Altres
La unitat de vida mínima
No coneixem del tot els detalls de què passa dins d’una cèl·lula. Per això la recerca intenta crear la unitat viva més simple possible, 35 cops més petita que la Escherichia Coli, amb només uns centenars de gens en lloc de 5.000. (New Yorker 7/3/2022).
It was by accident that Antoni van Leeuwenhoek, a Dutch cloth merchant, first saw a living cell. He’d begun making magnifying lenses at home, perhaps to better judge the quality of his cloth. One day, out of curiosity, he held one up to a drop of lake water. He saw that the drop was teeming with numberless tiny animals. These animalcules, as he called them, were everywhere he looked—in the stuff between his teeth, in soil, in food gone bad. A decade earlier, in 1665, an Englishman named Robert Hooke had examined cork through a lens; he’d found structures that he called “cells,” and the name had stuck. Van Leeuwenhoek seemed to see an even more striking view: his cells moved with apparent purpose. No one believed him when he told people what he’d discovered, and he had to ask local bigwigs—the town priest, a notary, a lawyer—to peer through his lenses and attest to what they saw.
Today, we take for granted that we are made of cells—liquidy sacs containing the Golgi apparatus, the endoplasmic reticulum, the nucleus. We accept that each of us was once a single cell, and that packed inside it was the means to build a whole body and maintain it throughout its life. “People ought to be walking around all day, all through their waking hours, calling to each other in endless wonderment, talking of nothing except that cell,” the physician Lewis Thomas wrote, in his book “The Medusa and the Snail.” But telescopes make more welcome gifts than microscopes. Somehow, most of us are not itching to explore the cellular cosmos. Today, although there’s still no microscope capable of showing everything that’s happening inside a living cell in real time, biologists grasp the strangeness of the zone, bigger than atoms but smaller than cells, in which the machinery of life exists. They’ve analyzed the tiny parts from which cells are made and learned how those parts interact. They’ve frozen cells, photographed them, and used computer simulations to revivify the pictures. They’ve studied the apparently empty spaces inside cells and discovered that they contain a world governed by unintuitive physical laws.
Several groups of “synthetic biologists” are now close to assembling living cells from nonliving parts. If we could design and control such cells with precision, we could use them to do what we want—generate clean energy, kill cancers, even reverse aging. The work depends on understanding a cell’s inner workings to a degree that van Leeuwenhoek could not have imagined.
They’ve modified a species of bacterium to create a “minimal” cell. It contains only what’s necessary for life—it’s the cellular equivalent of a stock car onto which new components can be bolted. John Glass, one of the project’s leaders, described the minimal cell to me as “a platform for figuring out the first principles in biology.”
J. Craig Venter, an instrumental player in efforts to sequence the human genome, felt a need to simplify. Why not create a cell with as few genes as possible, and use it as a model organism? If you wanted to understand a more complicated biological process, you could add the genes for it to your minimal cell. Venter assembled a team of biologists that included Glass, who was one of the world’s leading experts on a bacterium called Mycoplasma. “If you went to the zoo and lined up all the mammals and swabbed their urogenital tracts, you would find that each of them has some mycoplasma,” Glass told me. Because the bacteria live in such a nutrient-rich environment, they rarely have to forage for food, or even do much to digest it;
By 2016, after a few revisions, they had devised a minimal Mycoplasma genome half the size of the original. A researcher named Carole Lartigue spent years during her postdoc solving the daunting problem of implanting the genome in a cell. The bacterium that eventually resulted from the work was called JCVI-syn3.0. It was an engine bolted to some wheels.
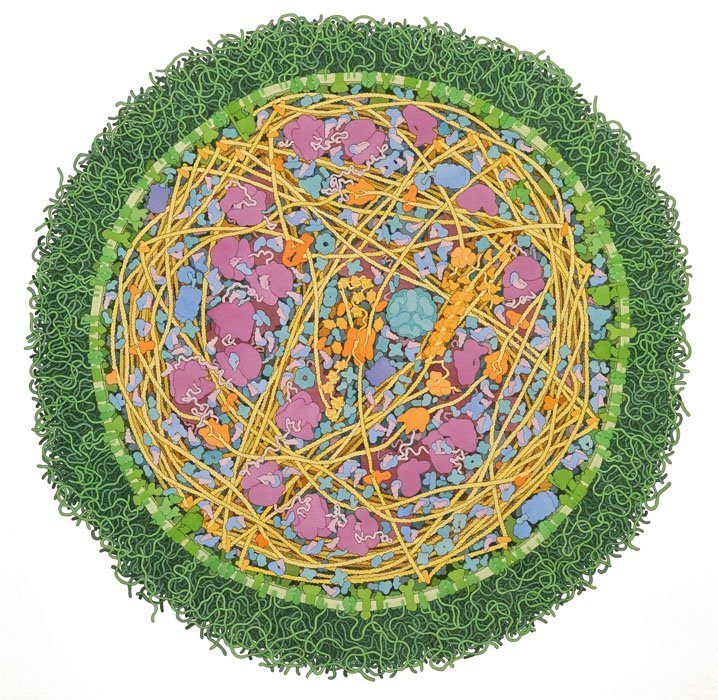
For contrast, Cook had prepared samples that contained both JCVI-syn3A and E. coli. The lab rat of biology, E. coli grows quickly and uniformly, and is genetically manipulable. It also hunts and eats, has a rudimentary kind of memory, and possesses around five thousand genes, compared with the minimal cell’s roughly five hundred. After Cook loaded the syn3A slide, I peered through the eyepiece, but struggled to distinguish the minimal cells from the floaters in my eyes. Then I looked at the other slide. An E. coli swam by. It was about thirty-five times bigger than the minimal cell by volume, and crenellated with complexity—a destroyer rather than a dinghy.
He showed me a poster noting all of JCVI-syn3A’s genes. About a third were labelled as having an unknown function. When the project began, there were a hundred and forty-nine mystery genes. Now about a hundred were left.
Generally, what a gene does depends on the protein it tells our cells to make. It’s proteins that run the cellular world, by sparking chemical reactions, sending signals, and self-assembling into biological machines. To understand and control a cell, or to design a new one, biologists need to know exactly how a given protein behaves in the cellular environment. What shapes can it take? What does it interact with? What happens when a small molecule, like a drug, gets lodged in one of its crevices?
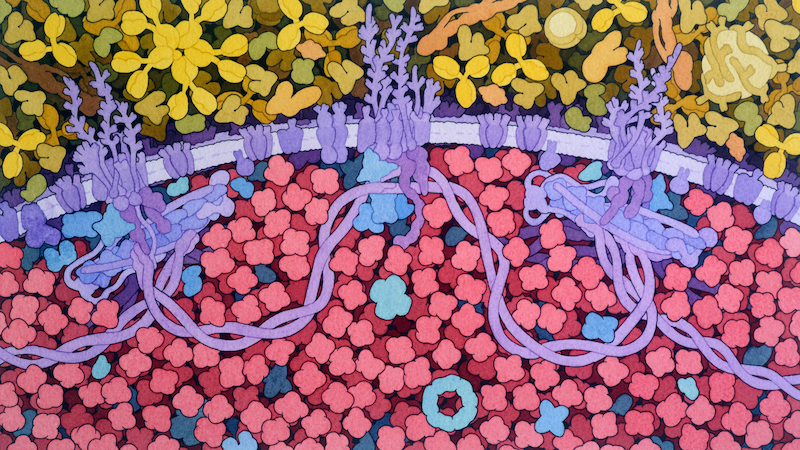
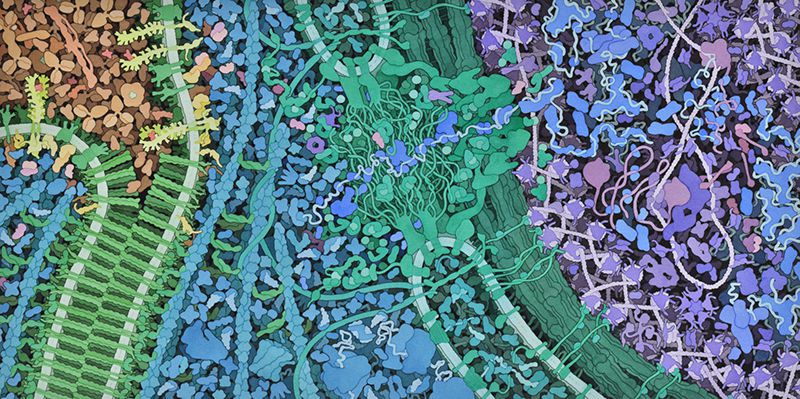
Our best pictures of the protein-rich cellular interior have come not from a microscope but from the brush of David S. Goodsell, a sixty-year-old biologist and watercolorist at the Scripps Research Institute. When I met Goodsell at Scripps, which is just down the road from J.C.V.I., he had long hair, a full beard, and a funky face mask. A painter since the age of ten, he illustrated his first E. coli during his postdoc, in 1991; the article that resulted, “Inside a Living Cell,
Roseanna N. Zia, a physicist who studies cells, emphasized the importance of physicality in biology. She told me that there were other “colloidal” properties of the cytoplasm, besides liquid-liquid phase separation, that nature might be using to its advantage—for instance, the fact that a shove at one end of the cytoplasm propagates, nearly instantly, to the other. Her group models how individual molecules subtly interact. “This area of understanding how colloidal-scale physics is regulating and orchestrating cell function—this is the frontier,” she said.
[ semblava que la biologia es reduïa a química i la química a física, i tot just estem aprenent a mirar les cèl·lules més simples!]
Cells, not DNA, are the master architects of life. No podem explicar encara què determina l’organisme a partir del DNA, sense l’entorn de la cèl·ula. No podem generar un ésser viu simplement a partir de la informació. I mlagrat les esperances inicials, tampoc podem posar una cèl·lula mare en una placa de Petri i desenvolupar el teixit que vulguem, cal un entorn determinat d’un conjunt de cèl·lules.